




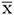 = 0.0215 s). La frecuencia de inicio más baja registrada fue de 322.7 Hz y la más alta de 13 956.5 Hz. El delta de frecuencias osciló entre 113.8 y 13 652.2 Hz., y el pico de frecuencias de 0 a 10 077.6 Hz. El 37.4 % de los sonidos emitidos por los titíes cabeciblancos carecen de armónicos y el 62.6 % restante presentan entre uno y doce armónicos.
= 0.0215 s). La frecuencia de inicio más baja registrada fue de 322.7 Hz y la más alta de 13 956.5 Hz. El delta de frecuencias osciló entre 113.8 y 13 652.2 Hz., y el pico de frecuencias de 0 a 10 077.6 Hz. El 37.4 % de los sonidos emitidos por los titíes cabeciblancos carecen de armónicos y el 62.6 % restante presentan entre uno y doce armónicos.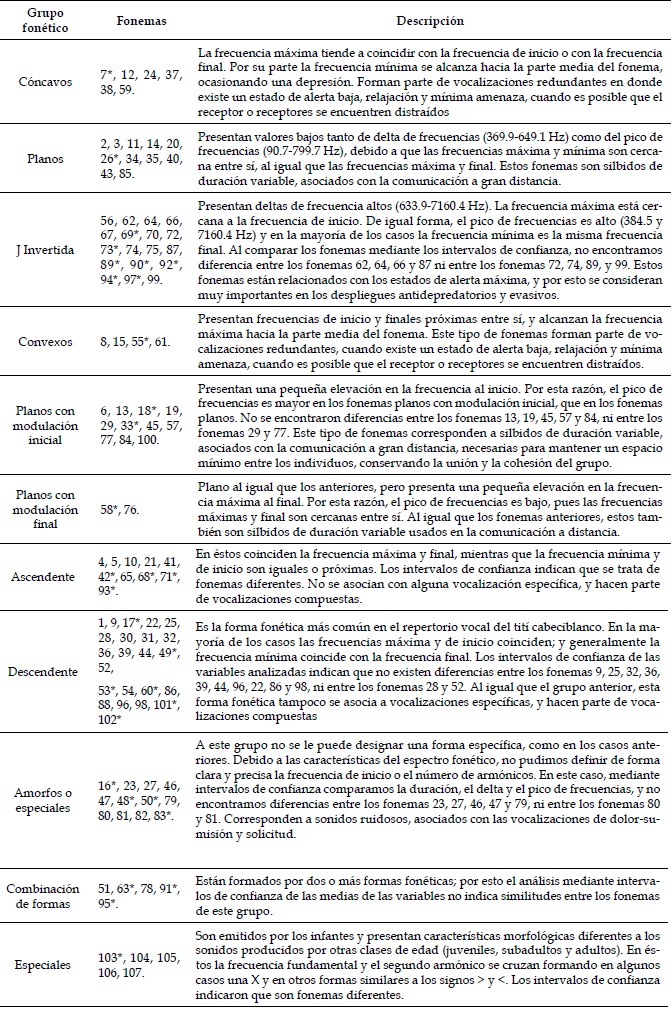

Recibido: 31 de mayo de 2020; Aceptado: 9 de noviembre de 2020
Resumen
Estudiamos titíes cabeciblancos (Saguinus oedipus, Primates: Callitrichidae) durante más de 100 horas en el municipio de Santa Catalina, Bolívar norte de Colombia. Grabamos 16 horas de sus vocalizaciones, las cuales digitalizamos a 44.1 kHz, utilizando Protools. Analizamos 2617 sonidos con Raven 1.2, con el fin de identificar las unidades fonéticas que conforman el repertorio vocal de esta especie, de acuerdo a los siguientes parámetros acústicos: duración (ms), delta de frecuencia (Hz), pico de frecuencias (Hz), frecuencia de inicio (Hz) y cantidad de armónicos de cada unidad fonética. Para describir el repertorio vocal, clasificamos los fonemas en 11 grupos, según su forma. Con el análisis de los intervalos de confianza de los parámetros acústicos, identificamos 49 sonidos diferentes. Mediante el ensamblaje de las unidades fonéticas, se identificaron 14 vocalizaciones diferentes. Los sonidos emitidos por el tití cabeciblanco fueron de corta duración (X̄ = 0.0215 s). La frecuencia de inicio más baja registrada fue de 322.7 Hz y la más alta de 13 956.5 Hz. El delta de frecuencias osciló entre 113.8 y 13 652.2 Hz., y el pico de frecuencias de 0 a 10 077.6 Hz. El 37.4 % de los sonidos emitidos por los titíes cabeciblancos carecen de armónicos y el 62.6 % restante presentan entre uno y doce armónicos.
Palabras clave:
Comunicación vocal, Etología, Primates, Secuencia fonética, Vocalizaciones..Abstract
We observed a population of cotton-top tamarins (Saguinus oedipus, Primates: Callitrichidae) in the municipality of Santa Catalina, Bolívar, northern Colombia. We observed them over 100 hours, and we recorded 16 hours of vocalizations, which were subsequently digitalized at 44.1 kHz, using Protools. We analyzed 2617 sounds with Raven 1.2, with the aim of identifying the phonetic units forming the vocal repertoire of this species, according to the following acoustic parameters: Duration (ms), change frequency (Hz), peak frequency (Hz), start frequency (Hz) and harmonics. In order to describe the vocal repertoire, the phonemes were classified into 11 groups, according to their form. The acoustic parameters were analyzed using confidence intervals, and 49 different sounds were identified. By assembling the phonetic units, 14 different vocalizations wer identified. The sounds emitted by cotton-top tamarins are of short duration (X̄ = 0.0215 s). The lowest start frequency registered is 322.7 Hz and the highest is 13 956.5 Hz. The change in frequency ranges from 113.8 to 13 652.2 Hz. The peak frequencies range from 0 to 10 077.6 Hz. On the other hand, 37.4 % of sound lacks harmonics, and the remainder 62.6 % show between 1 and 12 harmonics.
Keywords:
Ethology, Primates, Phonetic secuency, Vocalizations, Vocal communication..Introducción
El tití cabeciblanco (Saguinus oedipus), una especie endémica del Caribe colombiano, fue declarada En Peligro Crítico de extinción (CR), tanto a nivel global (Savage & Causado, 2014) como a nivel nacional (Ministerio de Ambiente y Desarrollo Sostenible, 2017), y se encuentra listada en el apéndice I de la CITES (Convención Internacional sobre el Comercio de Especies Amenazadas de Fauna y Flora).
Las vocalizaciones del tití cabeciblanco han sido estudiadas en cautiverio (Cleveland & Snowdon, 1982; Elowson et al., 1991; Roush & Snowdon, 1999; Castro & Snowdon, 2000; Weiss & Hauser, 2002; Miller et al., 2004; Miller et al., 2005; Campbell & Snowdon, 2007; 2009), pero se desconocen en vida silvestre. Cleveland & Snowdon (1982) describieron el repertorio vocal de un tití cabeciblanco adulto, utilizando una colonia de animales mantenidos en cautiverio en la Universidad de Wisconsin, Madison (Estados Unidos). Describieron la secuencia de los llamados, afirmando que el repertorio vocal de esta especie está derivado de dos elementos básicos: los llamados cortos de frecuencia modulada, chirps (que presentan ocho variantes fonéticas) y los llamados largos de frecuencia constante, whistles (con cinco variantes), y la combinación secuencial de estos elementos.
Cleveland & Snowdon (1982) identificaron 38 tipos diferentes de sonidos o combinaciones de sonidos. Sin embargo, es posible que existan variaciones en las características acústicas de los llamados en vida silvestre, puesto que los animales están inmersos en un ambiente sonoro, expuestos a depredadores, y con la necesidad de coordinar actividades como la alimentación, los desplazamientos y la vigilancia. Tales factores podrían modificar la estructura y la función de los llamados. Además, es importante resaltar que en cautiverio no se dieron encuentros agresivos intergrupales por la defensa del territorio (Cleveland & Snowdon, 1982); por esta razón es probable que en vida silvestre el repertorio de vocalizaciones sea diferente al observado en cautiverio.
En este estudio, describimos las unidades fonéticas que conforman el repertorio vocal del tití cabeciblanco en vida silvestre, teniendo en cuenta sus características acústicas, duración, delta de frecuencias, frecuencia de inicio, pico de frecuencias, número de armónicos y forma de cada elemento. Posteriormente, se analizó el ensamblaje de estas unidades fonéticas en vocalizaciones, teniendo en cuenta la actividad realizada por el emisor y el contexto en el que se presentó la vocalización. El presente estudio es el primer aporte sobre las vocalizaciones del tití cabeciblanco en vida silvestre y el primero en describir el repertorio vocal de los infantes.
Materiales y métodos
Realizamos el seguimiento, observación y grabación de dos grupos de titíes cabeciblancos silvestres, conformados en total por 15 individuos de todas las clases de edad, sexo y posición jerárquica, en un remanente de bosque seco tropical del municipio de Santa Catalina, Bolívar (10º 37’ 36” N-75° 14’50” O, 34 m s.n.m.). Los grupos empleados en este estudio se encuentran habituados a la presencia de investigadores en el área, debido a que fueron seguidos y observados diariamente por el equipo de la Fundación Proyecto Tití en sus investigaciones de campo. Realizamos las observaciones de los individuos con el método de animal focal y registro continuo (Altmann, 1974). La fase de campo fue de cuatro meses, con períodos de muestreo de quince días cada mes. Grabamos las vocalizaciones con una grabadora Sony TCM5000EV con respuesta de frecuencias de 20-20 000 Hz y frecuencia de muestreo de 44.1 KHz. Usamos 20 cassettes Quantegy cromados de 60 minutos de duración, un micrófono unidireccional Sennheiser M67 con respuesta a frecuencias entre 50 y 20 000 Hz; pantalla contra el viento Sennheiser MZW67, fuente de poder Sennheiser K6, soporte para el micrófono Audiotécnica AT8415, y cable para el micrófono. Para minimizar el ruido, por movimientos del investigador, utilizamos un soporte de madera de 1 m de largo, con un extremo puntiagudo enterrado en el suelo; y el otro extremo con una horqueta en la que se ubicó el micrófono dirigido hacia el animal focal.
Señalamos en la grabación el nombre de los individuos observados, la descripción de la vocalización, la identificación del emisor, la actividad realizada y el tipo de respuesta obtenida. Los individuos fueron previamente capturados y marcados con tinturas de colores en las partes blancas de su pelaje, para facilitar la identificación individual del emisor. Adicionalmente anotamos la hora, la fecha, las características climáticas, la presencia de grupos vecinos, el alimento consumido, el estrato del bosque y la presencia de otras especies. Grabamos siempre que se presentó un despliegue de vocalizaciones.
Digitalizamos las grabaciones utilizando Protools 5.1® en el Banco de Sonidos Animales del Instituto Alexander von Humboldt (BSA-IAvH), hoy Colección de Sonidos Ambientales, en formato AIFF. Una vez adquiridos, analizamos los espectrogramas utilizando el software Raven Sound Analysis versión 1.2 para Windows (Frecuencia: longitud DFT = 512 muestras, ventana Hann, ancho de banda = 124 Hz, resolución = 86.1 Hz; Tiempo: resolución = 5.8 ms).
Seleccionamos segmentos sin interferencias, por ruido antrópico o natural, en los que medimos sobre la frecuencia fundamental los siguientes parámetros acústicos: 1. Tiempo de inicio (ms), que corresponde al punto en el eje X en el que comienza la señal acústica; 2. Tiempo final (ms), que corresponde al punto en el eje X en el que termina la señal acústica. 3. Frecuencia de inicio (Hz), que corresponde al punto en el eje Y en el que inicia la señal. 4. Frecuencia final (Hz), como el punto en el eje Y en el que finaliza la señal. 5. Frecuencia mínima (Hz), como el punto en el eje Y que corresponde a la frecuencia más baja de la señal acústica. 6. Frecuencia máxima (Hz), como el punto en el eje Y que corresponde a la frecuencia más alta de la señal. 7. Número de armónicos, como la cantidad de frecuencias por encima o múltiplos de la frecuencia fundamental.
Para analizar los sonidos emitidos por el tití cabeciblanco, a partir de las variables anteriores derivamos las siguientes variables: Duración de la señal (ms), obtenida como la diferencia entre el tiempo final y el tiempo de inicio; delta de frecuencias (Hz), como la diferencia entre la frecuencia máxima y la frecuencia mínima; y el pico de frecuencias (Hz), obtenido como la diferencia entre la frecuencia máxima y la frecuencia final de cada fonema.
No se alteraron las características acústicas de los sonidos, puesto que no utilizamos filtros para mejorar la calidad auditiva de las grabaciones en ninguna parte del proceso de adquisición o análisis. Identificamos cada cassette o cinta de grabación con un acrónimo que corresponde a las iniciales del colector, seguido por la fecha de grabación, según las sugerencias del BSA-IAvH, entidad que es el depositario final de las grabaciones originales.
Realizamos pruebas para comprobar los supuestos de normalidad, aleatoriedad y homocedasticidad, respectivamente, en las variables medidas a cada fonema (Sokal & Rolhf, 1995). Para determinar el número de fonemas diferentes que conforman el repertorio vocal y el número de vocalizaciones de este primate optamos por un análisis descriptivo, comparando intervalos de confianza de las medias de las variables en estudio al 95 % de confianza, ya que son de gran utilidad porque dan estimaciones puntuales y brindan información sobre la precisión (Cumming & Finch, 2005)
Para describir el repertorio vocal, clasificamos los fonemas en 11 grupos según su forma, ocasionada por los cambios de frecuencia a lo largo del tiempo. Construimos y comparamos los intervalos de confianza al interior de cada grupo morfológico para las medias de las variables de duración, delta de frecuencias, frecuencia de inicio, pico de frecuencias y número de armónicos. Además, describimos el comportamiento de los animales al momento de emitir una vocalización específica. Los nombres dados en este estudio a algunas vocalizaciones tienen su origen en la nominación y descripción dada a las mismas por el equipo de la Fundación Proyecto Tití, en sus observaciones diarias del comportamiento de los animales.
Resultados
Observamos los titíes por más de 100 horas, en las que grabamos 16 horas de vocalizaciones. De éstas, medimos y analizamos 2617 sonidos agrupados en 107 fonemas. Por otra parte, analizamos el ensamblaje de estos fonemas en vocalizaciones, identificando 14 tipos de vocalizaciones de acuerdo con sus características auditivas, la actividad realizada y el comportamiento de los animales.
Los sonidos emitidos por el tití cabeciblanco fueron de corta duración. La duración mínima fue de 0.01 s; frente a una duración máxima de 3.438 s. Aproximadamente el 88 % de los sonidos duraron menos de 0.5 s, y cerca del 11 % de ellos tuvieron una duración de 0.5-1 s, mientras que muy pocos sonidos (1 %) presentaron una duración mayor a 1 s.
La frecuencia más baja de inicio de vocalizaciones fue 322.7 Hz y la más alta fue 13 956.5 Hz. Las frecuencias de inicio más empleadas por esta especie (34 %), se extienden en el rango 2692.31 - 3923.08 Hz. El delta de frecuencias (frecuencia máxima-frecuencia mínima) fue entre 113.8 y 13 652.2 Hz. El 58 % de los sonidos presentaron un delta de frecuencias entre 239.8 y 1461.5 Hz, mientras que solo el 1 % presentan un delta de frecuencias mayor a 10 kHz. El pico de frecuencias (diferencia entre frecuencia máxima y frecuencia final de un sonido) estuvo entre 0 y 10 077.6 Hz. Mientras que cerca del 60 % de los sonidos emitidos por esta especie tuvieron un pico de frecuencias entre 0 y 846.154 Hz.
Las medias (X̄ ), desviaciones estándar (s) y los tamaños de muestra de cada variable analizada se presentan en la Tabla 1. De otra parte, el 37.4 % de los sonidos emitidos por este primate carecen de armónicos, mientras que el 62.6 % restante tienen entre 1 y 12 armónicos.
* Fonemas excluidos del análisis porque no tienen el tamaño mínimo de muestra
Tabla 1: Descripción de unidades fonéticas del repertorio vocal del tití cabeciblanco, Saguinus oedipus, en Santa Catalina, Bolívar, norte de Colombia.
Grupo fonético
Fonemas
Descripción
Cóncavos
7*, 12, 24, 37, 38, 59.
La frecuencia máxima tiende a coincidir con la frecuencia de inicio o con la frecuencia final. Por su parte la frecuencia mínima se alcanza hacia la parte media del fonema, ocasionando una depresión. Forman parte de vocalizaciones redundantes en donde existe un estado de alerta baja, relajación y mínima amenaza, cuando es posible que el receptor o receptores se encuentren distraídos
Planos
2, 3, 11, 14, 20, 26*, 34, 35, 40, 43, 85.
Presentan valores bajos tanto de delta de frecuencias (369.9-649.1 Hz) como del pico de frecuencias (90.7-799.7 Hz), debido a que las frecuencias máxima y mínima son cercana entre sí, al igual que las frecuencias máxima y final. Estos fonemas son silbidos de duración variable, asociados con la comunicación a gran distancia.
J Invertida
56, 62, 64, 66, 67, 69*, 70, 72, 73*, 74, 75, 87, 89*, 90*, 92*, 94*, 97*, 99.
Presentan deltas de frecuencia altos (633.9-7160.4 Hz). La frecuencia máxima está cercana a la frecuencia de inicio. De igual forma, el pico de frecuencias es alto (384.5 y 7160.4 Hz) y en la mayoría de los casos la frecuencia mínima es la misma frecuencia final. Al comparar los fonemas mediante los intervalos de confianza, no encontramos diferencia entre los fonemas 62, 64, 66 y 87 ni entre los fonemas 72, 74, 89, y 99. Estos fonemas están relacionados con los estados de alerta máxima, y por esto se consideran muy importantes en los despliegues antidepredatorios y evasivos.
Convexos
8, 15, 55*, 61.
Presentan frecuencias de inicio y finales próximas entre sí, y alcanzan la frecuencia máxima hacia la parte media del fonema. Este tipo de fonemas forman parte de vocalizaciones redundantes, cuando existe un estado de alerta baja, relajación y mínima amenaza, cuando es posible que el receptor o receptores se encuentren distraídos.
Planos con modulación inicial
6, 13, 18*, 19, 29, 33*, 45, 57, 77, 84, 100.
Presentan una pequeña elevación en la frecuencia al inicio. Por esta razón, el pico de frecuencias es mayor en los fonemas planos con modulación inicial, que en los fonemas planos. No se encontraron diferencias entre los fonemas 13, 19, 45, 57 y 84, ni entre los fonemas 29 y 77. Este tipo de fonemas corresponden a silbidos de duración variable, asociados con la comunicación a gran distancia, necesarias para mantener un espacio mínimo entre los individuos, conservando la unión y la cohesión del grupo.
Planos con modulación final
58*, 76.
Plano al igual que los anteriores, pero presenta una pequeña elevación en la frecuencia máxima al final. Por esta razón, el pico de frecuencias es bajo, pues las frecuencias máximas y final son cercanas entre sí. Al igual que los fonemas anteriores, estos también son silbidos de duración variable usados en la comunicación a distancia.
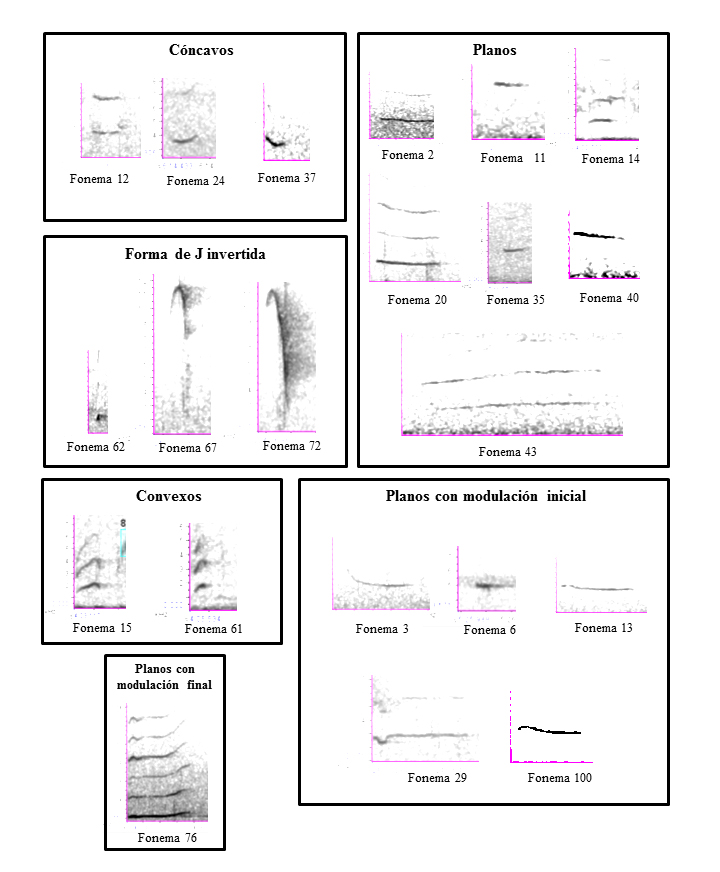
Ascendente
4, 5, 10, 21, 41, 42*, 65, 68*, 71*, 93*.
En éstos coinciden la frecuencia máxima y final, mientras que la frecuencia mínima y de inicio son iguales o próximas. Los intervalos de confianza indican que se trata de fonemas diferentes. No se asocian con alguna vocalización específica, y hacen parte de vocalizaciones compuestas.
Descendente
1, 9, 17*, 22, 25, 28, 30, 31, 32, 36, 39, 44, 49*, 52, 53*, 54, 60*, 86, 88, 96, 98, 101*, 102*
Es la forma fonética más común en el repertorio vocal del tití cabeciblanco. En la mayoría de los casos las frecuencias máxima y de inicio coinciden; y generalmente la frecuencia mínima coincide con la frecuencia final. Los intervalos de confianza de las variables analizadas indican que no existen diferencias entre los fonemas 9, 25, 32, 36, 39, 44, 96, 22, 86 y 98, ni entre los fonemas 28 y 52. Al igual que el grupo anterior, esta forma fonética tampoco se asocia a vocalizaciones específicas, y hacen parte de vocalizaciones compuestas
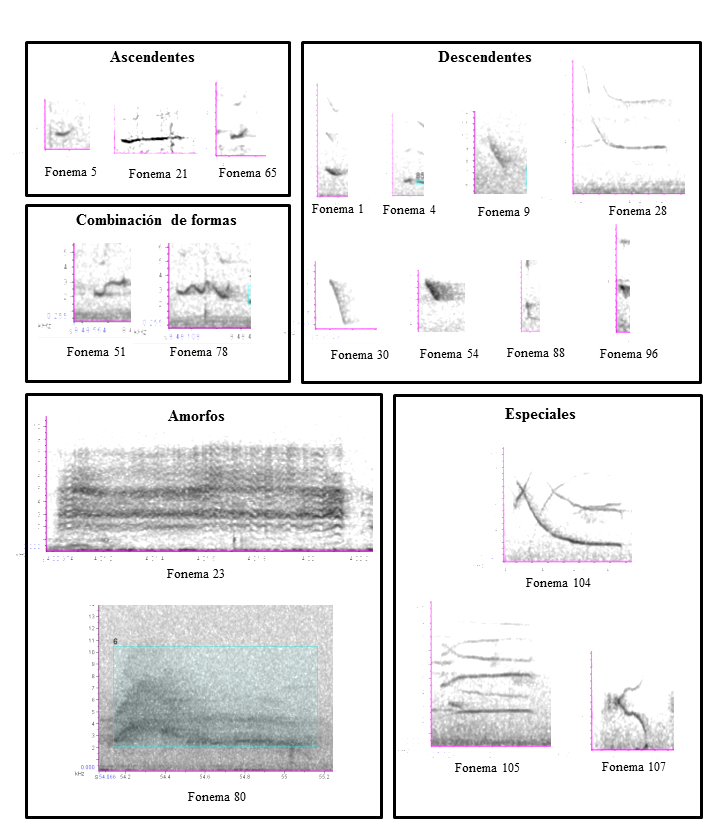
Amorfos o especiales
16*, 23, 27, 46, 47, 48*, 50*, 79, 80, 81, 82, 83*.
A este grupo no se le puede designar una forma específica, como en los casos anteriores. Debido a las características del espectro fonético, no pudimos definir de forma clara y precisa la frecuencia de inicio o el número de armónicos. En este caso, mediante intervalos de confianza comparamos la duración, el delta y el pico de frecuencias, y no encontramos diferencias entre los fonemas 23, 27, 46, 47 y 79, ni entre los fonemas 80 y 81. Corresponden a sonidos ruidosos, asociados con las vocalizaciones de dolor-sumisión y solicitud.
Combinación de formas
51, 63*, 78, 91*, 95*.
Están formados por dos o más formas fonéticas; por esto el análisis mediante intervalos de confianza de las medias de las variables no indica similitudes entre los fonemas de este grupo.
Especiales
103*, 104, 105, 106, 107.
Son emitidos por los infantes y presentan características morfológicas diferentes a los sonidos producidos por otras clases de edad (juveniles, subadultos y adultos). En éstos la frecuencia fundamental y el segundo armónico se cruzan formando en algunos casos una X y en otros formas similares a los signos > y <. Los intervalos de confianza indicaron que son fonemas diferentes.
Aunque con base en estas variables identificamos 107 fonemas, para este análisis descartamos 31 fonemas con menos de 10 datos ya que el intervalo de confianza resultante fue demasiado amplio. Algunos fonemas son iguales entre sí y en este caso los consideramos como uno sólo. Por lo tanto, el repertorio vocal de este primate tuvo aproximadamente entre 49 y 80 unidades fonéticas, que se agruparon en 11 formas fonéticas. La descripción de los grupos fonéticos se presenta en la Tabla 1 y se ilustran en la Figura 1.
Figura 1: Grupos fonéticos del tití cabeciblanco (Saguinus oedipus) silvestre en el bosque seco tropical de Santa Catalina (Bolívar), Colombia, organizados según su morfología.
Tabla 2: Descripción de las vocalizaciones del tití cabeciblanco (Saguinus oedipus) silvestre en el bosque seco tropical de Santa Catalina (Bolívar), Colombia.
Vocalización
Descripción acústica
Comportamiento
Actividad /Contexto
Agresiva
Vocalización compuesta. En esta aparecen los fonemas 3, 12, 15, 27, 28, 37, 38, 39, 44, 61, 62, 64, 65, 66, 67, 70, 72, 74, 75, 76, 77, 78 y 87, los cuales corresponden principalmente a fonemas con forma de J invertida. El 64.3 % de las vocalizaciones iniciaron con el fonema 72, que a su vez se encuentra en el 78.6 % del total analizado. Por su parte el fonema 74 aparece en el 85.7 %, siendo los fonemas característicos de esta vocalización. Estos fonemas tienen picos y deltas de frecuencia muy altos y presentan entre 0 y 6 armónicos, siendo más frecuente un armónico. Es una vocalización principalmente aguda, FMáx= 13.9 kHz., y de larga duración (X±s ± 4.21 ± 2.48 s),
El despliegue de vocalizaciones agresivas puede registrarse durante varios minutos, tanto como permanezca en el área el agente que lo originó. Se emite con la boca visiblemente abierta, usualmente va acompañada de piloerección en la cola y/o cabeza, exhibición corporal, excitación y movimientos de la lengua. El emisor puede desplazarse horizontal y verticalmente en el árbol con movimiento errático o sólo mover la cabeza sin cambiar su posición en el mismo.
El despliegue agresivo generalmente es iniciado por el individuo que está realizando la actividad de vigilancia dentro del grupo y puede ser imitado por otros integrantes. No obstante, algunos de ellos pueden emitir otros tipos de vocalizaciones, que incluyen vocalización de exploración, nerviosa, pelea o, incluso, solicitud, dependiendo del grado de alerta en que se encuentre cada uno de ellos y la clase de edad. La vocalización agresiva puede presentarse como respuesta a un estímulo repentino o a la permanencia en el sitio de una fuente potencial de peligro. Hace parte del comportamiento antidepredatorio de la especie, pues en este caso, los individuos adultos y subadultos emiten esta vocalización con el fin de ahuyentar o disuadir al posible depredador. En algunas ocasiones esta vocalización fue dirigida hacia los observadores; en otros casos estuvo relacionada con estímulos del medio.
Alarma
Es una vocalización aguda (X
fmáx
± s = 9243.7 ± 741,25 Hz), alcanza una frecuencia máxima de 13.9 kHz; de muy corta duración (X ± s = 0.324 ± 0.131s). Está conformada por el fonema 99, que se repite dos veces. La vocalización de alarma carece de armónicos.
Se emite con la boca abierta y es producida por quien realiza la actividad de vigilancia. Como respuesta, los individuos del grupo reaccionan de manera inmediata, manifestando un evidente incremento en el estado de alerta general.
Está asociada a la cercanía de un potencial depredador o peligro; fue observada en presencia principalmente de aves rapaces como Milvago chimachima y Rupornis magnirostris. También fue registrada ante Catharthes aura. Un infante debido a su inexperiencia emitió vocalización de alarma ante la presencia de una ardilla (Sciurus granatensis).
Alimentación
Es una vocalización de corta duración (X±s = 0.56 ± 0,83 s). Presenta la secuencia de fonemas 30, 9, 25.
Producida con la boca semiabierta; puede ser emitida por cualquier individuo.
Esta vocalización se registró cuando los animales encontraron fuentes de alimento y fue emitida mientras se manipulaba el fruto hallado.
Contacto
Es la vocalización más frecuente (222) y representa el 51.1 % de las vocalizaciones registradas y analizadas. Por ser una vocalización simple, es muy corta (X± s ± 0.44 ± 0.18 s). Se registraron 11 fonemas diferentes para esta vocalización: 13, 20, 28, 29, 40, 43, 45, 57, 77, 84, y 100; no obstante, los más representativos son los fonemas 13 (32.8 %), 57 (17.1 %), 28 (13 %), 100 (9.9 %) y 20 (9 %), los cuales corresponden a silbidos largos. Los armónicos se presentan en número variable.
Producida con la boca semiabierta; puede ser emitida por cualquier individuo del grupo generalmente cuando se encuentran dispersos (aun en el mismo árbol) y se presenta en diferentes actividades.
Se presenta en medio de despliegues de vocalizaciones de exploración, pelea, agresiva, nerviosa, vigilancia y solicitud. La respuesta que evoca puede ser otra vocalización de contacto o vocalizaciones de llamado.
Dolor-Sumisión
Es una vocalización con una duración intermedia (X ± s = 1.64 ± 0.95 s). En algunos casos se presentó como vocalización compuesta, en otros como vocalizaciones simples. Los fonemas con que se representa son amorfos; incluye los fonemas 23, 79, 80 y 81. Los más representativos son los fonemas 23 y 79 (76.9 %)
Se trata de un grito agudo, emitido con la boca abierta. Es emitida por el receptor de una agresión física. Generalmente va acompañada de intento de huida y de otras vocalizaciones más suaves que tienen como fin apaciguar la situación y evitar la agresión.
Se presenta en los encuentros intergrupales, cuando un individuo aleja a la hembra dominante de su grupo del sitio donde se está dando el enfrentamiento, con el fin de evitar que se acerque a los machos del grupo contrario. También se presenta en los encuentros agresivos intragrupales, cuando un individuo dominante rechazó una oferta de acicalamiento por parte de un subordinado; y cuando un subordinado no respetó las jerarquías en un evento de alimentación. Adicionalmente, se registró durante el juego entre infantes y/o juveniles.
Exploración
Es una vocalización de frecuencia media (alrededor de 3 kHz), de duración intermedia (X ± s = 1.64 ± 0.56 s), representada por fonemas descendentes, planos, ascendentes, cóncavos, convexos y con forma de J invertida, reuniendo los fonemas 1, 2, 3, 4, 10, 11, 12, 21, 24, 25, 36, 37, 39, 61 y 66. Los elementos constitutivos más importantes de esta vocalización son los fonemas 3 (13.8%) y 37 (43.6 %). En general, los fonemas en esta vocalización presentan un número variable de armónicos. No se define una secuencia clara en esta vocalización, pues al parecer los elementos podrían ser pronunciados de manera independiente, con redundancia.
Puede ser emitida con la boca semiabierta, por cualquier individuo del grupo, en diferentes actividades y situaciones.
Presente en todas las actividades que realizan estos primates. Aparece en periodos de tranquilidad, acompañada de un estado de alerta mínimo. Es producida durante el forrajeo, descanso y actividades sociales (acicalamiento, juegos y encuentros agresivos). En un despliegue agresivo, también puede presentarse vocalización de exploración por un individuo con un estado de alerta bajo.
Llamado
Es una vocalización larga (X ± s = 2.08 ± 0.15 s); presenta la secuencia de fonemas 84-85-85; y 28-85-85-85, variación relacionada al parecer con la actividad desarrollada, pues registramos la segunda secuencia en los encuentros agresivos intergrupales por la defensa del territorio. En el caso de los infantes, la vocalización de llamado está compuesta por la secuencia 104-105-105.
Esta vocalización es producida con la boca abierta. Puede ser emitida por cualquier miembro del grupo; generalmente la respuesta que evoca es otra vocalización de llamado o de contacto.
Grabada durante diferentes actividades; es muy importante antes, durante y después de los encuentros agresivos intergrupales por la defensa del territorio. También la registramos cuando el grupo se encontraba disperso y cuando los infantes requirieron apoyo de un individuo adulto en un desplazamiento.
Nerviosa
De larga duración (X ± s = 2.84 ± 0.54 s) y frecuencia media (2600-2800 Hz). Presenta los fonemas 1, 2, 3, 4, 6, 8, 13, 37, 38, 39, 40, 41, 42, 43, 44, 56, 67, 74, 77 y 78; reúne ocho formas fonéticas. Los más representativos son los fonemas 38 (29.4%) y 39 (30%)
Es producida con la boca abierta, acompañada de movimientos rápidos; se presenta también algo de piloerección y exhibición del cuerpo.
Emitida ante un estímulo repentino; generalmente se presenta después de una vocalización de alarma, asociada a un alto grado de alerta. El despliegue nervioso puede durar tanto tiempo como permanezca el estímulo en el ambiente. En ocasiones es el preámbulo de un despliegue de vocalizaciones agresivas.
Pelea
Es una vocalización de corta duración (X ± s = 0.96 ± 0.31s). Agrupa los fonemas 11, 14, 15, 25, 28, 35, 64, 65, 66, 70, 72, 74, 85, 87, 88 y 96. Se presentaron cuatro vocalizaciones relacionadas con los encuentros intergrupales. La vocalización de pelea 1 tiene la secuencia 96-88-88-88. La vocalización de pelea 2 presenta la secuencia 101-89-89-89- 89-89. Por otra parte la vocalización de pelea 3 tiene la secuencia 14-14-14-14-14-14-14-14. Estas vocalizaciones son exclusivas de los encuentros agresivos intergrupales, mientras que la vocalización de pelea 4, con la secuencia 87-87-88-88- 88-88-88-88-88-88-88-88-88-88-87-87, se registró durante los despliegues agresivos ante un potencial peligro.
Van acompañadas de piloerección en la cabeza y cola, exhibición del cuerpo, excitación y movimientos de la lengua.
Se presentan en los encuentros agresivos intergrupales por la defensa del territorio, cuando hay contacto visual entre los grupos. Las vocalizaciones de pelea 1 y 2 fueron emitidas en presencia del grupo contrario; la vocalización de pelea 3 fue emitida por los individuos adultos antes del contacto físico. Los encuentros intergrupales incluyen persecuciones y agresión física, principalmente entre los machos dominantes. En los encuentros en los que participaron solo los subadultos no hubo agresión física, sino despliegues de vocalizaciones de pelea, llamado, contacto, agresiva y nerviosa. Los infantes y juveniles no participaron nunca de estos encuentros, pudiendo permanecer en una rama individuos de ambos grupos, vocalizando llamado, nerviosa y solicitud.
Solicitud
Es una vocalización de duración media (X ± s = 1.45 ± 0.47 s). Reúne los fonemas 6, 13, 23, 27, 28, 37, 39, 45, 47, 57, 77, 81, 85, 86, 104, 105, y 107. Los fonemas más representativos son el 23 (21.9 %), 104 (20.7%) y 107 (15.8%). La vocalización de solicitud de las crías sigue la secuencia 104-107-107, mientras en los juveniles es 23-23-23.
Va acompañada de vocalizaciones de contacto y de llamado y se produce con la boca semiabierta.
Es emitida principalmente por los infantes cuando requieren apoyo de un adulto o alimento, y en ocasiones por los juveniles. Los individuos adultos se acercan a los infantes y satisfacen esta necesidad, ya sea acarreándolos o proveyéndolos de alimento. Se presentó durante los encuentros agresivos intergrupales, en los desplazamientos del grupo y en los eventos de alimentación.
Vigilancia
Es una vocalización larga (X ± s = 4.18 ± 2.88 s). Al igual que en las vocalizaciones de exploración y nerviosa, en ésta son unos pocos elementos que se repiten varias veces en una misma vocalización Comprende los fonemas 1, 2, 3, 12, 14, 20, 35, 37, 38, 39, 40, 42, 43, 64, 77, 87, 88 y 100. Siendo los fonemas 37 (43.1%), 17 (16.6%) y 40 (9.8%) los más importantes.
Es un silbido, asociado a un estado de alerta baja.
Esta vocalización es emitida por quien realiza la actividad de vigilancia al interior del grupo.
Discusión
Los sonidos producidos por el tití cabeciblanco son de corta duración y principalmente agudos. Esto coincide con lo reportado por Morton (1977), quien afirma que en mamíferos y aves la frecuencia fundamental está negativamente correlacionada con el tamaño corporal. Esta afirmación ha sido corroborada por Hauser (1993); por su parte Moynihan (1970 citado por Oppenheimer, 1997) sugiere que los primates pequeños, al tener un riesgo de depredación más alto que los primates de mayor tamaño, han seleccionado frecuencias altas en su comunicación vocal, pues es más difícil ubicar un animal que emplea tales frecuencias. Este es el caso del tití cabeciblanco cuyas vocalizaciones en vida silvestre son principalmente agudas, posiblemente por su tamaño pequeño, y de corta duración, con el fin de minimizar el riesgo de depredación. Esto mismo ha sido reportado para esta especie en condiciones de cautiverio (Cleveland & Snowdon 1982; Campbell & Snowdon, 2007). También es similar a lo reportado para otras especies del género en estado silvestre en Colombia (Rueda & Zerda, 2009; Fuentes, 2006; Fuentes et al., 2013). Las vocalizaciones del tití cabeciblanco también presentan algunas características similares, en cuanto a su duración y forma, a las encontradas por Rueda & Zerda (2009), Fuentes (2006) y Fuentes et al.(2013), para el tití gris (Saguinus leucopus). Sin embargo, el tití cabeciblanco alcanza frecuencias más altas en sus vocalizaciones que las reportadas por estos autores para el tití gris en estado silvestre en un bosque de Mariquita, Tolima.
Cleveland & Snowdon (1982) describieron 38 sonidos o combinaciones de sonidos en el repertorio vocal de titíes cabeciblancos adultos mantenidos en cautiverio. En el presente estudio se identificaron 49 sonidos diferentes para la especie en estado silvestre, incluyendo la descripción de los sonidos o fonemas emitidos por infantes, que difieren significativamente de los emitidos por otras clases de edad, constituyéndose en el primer aporte sobre sonidos de los infantes y, en general, de esta especie endémica en condiciones naturales. En cautiverio, los infantes de titíes cabeciblancos producen sonidos que no se presentan en los adultos, aunque parecen comprender los sonidos emitidos por estos (Castro & Snowdon, 2000). Según Rueda & Zerda (2009) es posible que los infantes de titíes emitan vocalizaciones poco definidas y repetitivas, diferentes a otras clases de edad, comparable a lo que ocurre en el lenguaje infantil de los humanos.
La vocalización agresiva registrada para el tití cabeciblanco en vida silvestre, emitida ante un peligro potencial, hace parte del comportamiento antidepredatorio de la especie. Esta equivale al trino de gorjeos altos identificado por Campbell & Snowdon (2007) para titíes cabeciblancos en cautiverio, con similar duración, forma y frecuencia de inicio de sus elementos constituyentes. Sin embargo, en vida silvestre, la frecuencia máxima alcanzada en estas vocalizaciones es más alta que la reportada para titíes cautivos. Ellos describen una secuencia de sonidos agudos y de corta duración (bark bout), con características similares a la vocalización de pelea 4 de nuestro estudio, que también fue registrada en los despliegues de vocalizaciones agresivas. Al igual que en el estudio de Campbell & Snowdon (2007), encontramos que al ser este un comportamiento social, se presentaron coros de otros individuos del grupo que emitían la misma vocalización o vocalizaciones diferentes, dependiendo de la clase de edad y del estado motivacional. Una vez el estímulo desaparece o disminuye la amenaza, estas vocalizaciones van siendo sustituidas por vocalizaciones nerviosas, pasando a un estado de vigilancia. Sin embargo, contrario a lo predicho por Marler (1955 citado por Campbell & Snowdon, 2007) y Bradbury & Veherencamp (1998), quienes afirman que las vocalizaciones antidepredatorias deberían ser de baja frecuencia para facilitar la ubicación del emisor y trabajar conjuntamente en el ahuyentamiento del depredador, las vocalizaciones agresivas del tití cabeciblanco fueron principalmente agudas. Esto también contradice lo descrito por Morton (1977), quien afirma que las vocalizaciones producidas en contextos hostiles son de baja frecuencia.
Rueda & Zerda (2009) reportaron para el tití gris un sonido con forma de “U” invertida, emitido cuando existen amenazas, pudiendo presentarse secuencias de 2-50 señales de este tipo; tal sonido presenta características similares a los fonemas constituyentes de la vocalización agresiva. Por su parte, Fuentes (2006) y Fuentes et al. (2013) reportaron un gorjeo “A” para la misma especie, con forma similar a algunos elementos de la vocalización agresiva de nuestro estudio; sin embargo difiere en la duración y frecuencia, siendo aquéllos más largos y de tono más bajo que los registrados para el tití cabeciblanco, pero con funciones similares.
La vocalización de alarma, por su parte, concuerda con lo expuesto por Marler (1955 citado por Campbell & Snowdon, 2007), Bradbury & Vehrencamp (1998) y Brown (1982 citado por Inglis & Shepherd, 1990), ya que se trata de una vocalización de tono alto y corta duración, para evitar que el depredador pueda ubicar al emisor. Esta vocalización puede preceder o no a la vocalización agresiva, dependiendo de si los animales desean evadir o ahuyentar al peligro potencial. El reconocimiento de los depredadores y peligros potenciales para la especie es un comportamiento adquirido debido a su experiencia en el medio. No observamos ataques o intentos de ataque por otro tipo de depredadores (mamíferos terrestres o serpientes), por lo que no se pudo constatar si las vocalizaciones de alarma variaban según el tipo de depredador de acuerdo con lo registrado para otras especies de primates por Seyfarth & Cheney (1990).
El gorjeo tipo E reportado por Cleveland & Snowdon (1982), tiene funciones de evasión según estos autores; su duración, forma y frecuencia son similares a la vocalización de alarma reportada en nuestro estudio. En el caso del tití gris, Rueda & Zerda (2009) describen un piup con funciones de alarma; sin embargo este no presenta similitud alguna con la vocalización de alarma del tití cabeciblanco. Tampoco hay similitud entre esta vocalización y la alarma presentada por Fuentes et al. (2013). Por consiguiente, existe especificidad en los sonidos de alarma de cada especie.
Para titíes en cautiverio, Cleveland & Snowdon (1982) reportan un gorjeo tipo A, similar al fonema 30 que encontramos en este estudio, pero de frecuencia más baja. También describen el gorjeo tipo C, similar al fonema 9 de la vocalización de alimentación. Ellos señalan que este sonido se presenta ante la aproximación a una fuente de alimento. Además describen un gorjeo tipo D, cuando el animal tiene en su posesión comida u objetos; sin embargo no se encontró similitud entre éste y alguno de los elementos constituyentes de la vocalización de alimentación. Rueda & Zerda (2009), Fuentes (2006) y Fuentes et al. (2013) presentan para el tití gris un silbido A, asociado a contextos alimentarios, con características de duración y frecuencia similares al fonema 30 de la vocalización de alimentación; sin embargo difieren en la forma.
La vocalización de contacto es una vocalización simple, representada por un silbido largo. Es muy importante en la comunicación vocal del tití cabeciblanco, y fue la que más registramos en el estudio. Esta vocalización es empleada para mantener la cohesión del grupo y un espacio mínimo entre los individuos al realizar diferentes actividades. Es útil en la comunicación a distancia, y puede ser empleada para comunicarse, aun cuando el receptor o receptores no son visibles para el emisor. Se presentó en la comunicación desde 1 m hasta más de 50 m. Esta vocalización presenta características similares de duración y frecuencia con los pequeños silbidos modulados inicialmente, pequeños silbidos modulados al final, silbidos planos y silbidos múltiples ascendentes descritos por Cleveland & Snowdon (1982) en diferentes contextos, pero siempre con la función de mantener el contacto entre los miembros del grupo. Fuentes (2006), presenta un chiih A como un sonido con funciones de contacto, con variaciones en la respuesta provocada, según el contexto en el que fue emitido por el tití gris; sin embargo, este sonido no tiene equivalente dentro de las vocalizaciones de contacto del tití cabeciblanco. No obstante, este autor describe un piiip como un sonido de alarma emitido en diferentes actividades, mientras los animales se esconden y se asoman o huyen de una amenaza. Tal sonido presenta características similares a la vocalización de contacto del tití cabeciblanco; entonces, al presentarse en medio del desarrollo de diferentes actividades, podría tener funciones de contacto para mantener la cohesión del grupo, en lugar de funciones de alarma.
La vocalización de dolor-sumisión es un chillido ruidoso, emitido por el receptor de una agresión física, que busca apaciguar la situación y evitar la agresión. Conforme a lo descrito por Morton (1977), es un sonido de baja frecuencia, presente en un comportamiento hostil.
Generalmente está asociada a los encuentros agresivos intergrupales por la defensa del territorio y al mantenimiento del orden jerárquico. Existe similitud en las características acústicas de esta vocalización y los graznidos y gritos reportados por Cleveland & Snowdon (1982) para titíes cautivos, presentando la misma función. También es similar al chillido A descrito por Rueda & Zerda (2003) y al chirrido de Fuentes et al. (2013) para el tití gris; sin embargo en estos últimos estudios no especifican si el emisor de la vocalización es el agresor o el agredido.
La vocalización de exploración es una vocalización asociada a un estado de alerta curiosa, en el que no se presentan amenazas para los animales. Esta vocalización, registrada en la mayoría de las actividades realizadas por los titíes, se relaciona con la exploración del territorio en busca de alimentos, refugios y marcaje del mismo. Es una vocalización redundante, es decir, que sus elementos constitutivos se repiten constantemente con el fin de darle mayor duración a la señal en el medio, dado que según la actividad realizada el receptor o receptores podrían encontrarse distraídos. Entonces, se considera que esta vocalización tiene como objetivo informar a los miembros del grupo la ubicación del emisor así como la actividad realizada. Sin embargo, siempre hay un individuo que emite vocalización de vigilancia para garantizar la seguridad del grupo. El gorjeo tipo B descrito por Cleveland & Snowdon (1982), emitido hacia observadores u objetos familiares, presenta similitudes con algunos elementos de la vocalización de exploración. Los sonidos asociados con la exploración del territorio, descritos por Rueda & Zerda (2009) y Fuentes et al. (2013) para el tití gris, difieren completamente en su forma y características acústicas a los emitidos por el tití cabeciblanco, indicando que esta vocalización es especie-específica, dado que es necesaria para mantener la unidad grupal en la coordinación y desarrollo de diferentes actividades.
La vocalización de llamado es una vocalización compuesta por silbidos largos, utilizada en la comunicación a distancias superiores de 100 m cuando el grupo se encontró disperso o ante la cercanía de un grupo vecino. Fue la vocalización que más se presentó durante los encuentros agresivos intergrupales, por lo que no sólo tendría la función de reunir al grupo cuando está disperso, sino que también interviene en el mantenimiento y defensa del territorio. Al respecto registramos diferencias en las variables acústicas analizadas, de acuerdo con la actividad realizada y según la clase de edad del emisor. Cleveland & Snowdon (1982), presentan un llamado largo normal y el gorjeo tipo f + silbido con características de forma, secuencia, duración y frecuencia similares a la vocalización de llamado de este estudio. Ellos indican que se presentan como respuesta a los llamados de otros grupos y durante el aislamiento de un individuo. También presentan un llamado largo combinado emitido durante el juego, aislamiento, cuando los animales son socialmente molestados o cuando el animal no está visible; sin embargo ésta es similar a la vocalización de llamado registrada antes de los encuentros agresivos intergrupales, por lo que en estado silvestre tendría la función de reunir y preparar al grupo para defender su territorio. Vocalizaciones de llamado similares al llamado largo silencioso de estos autores, se registraron durante los encuentros intergrupos. Las vocalizaciones de llamado del tití cabeciblanco descritas en nuestro estudio difieren en su forma y características acústicas a los sonidos con la función de reunir al grupo en el tití gris descritos por Rueda & Zerda (2009); Fuentes (2006) y Fuentes et al. (2013) pero presentan similitudes con los llamados intergrupales de Saguinus geoffroyi descritos por Oppenheimer (1977).
La vocalización nerviosa, emitida ante un estímulo del medio que ofrece cierto grado de amenaza, es redundante al igual que la vocalización de exploración, para garantizar que todos los miembros del grupo estén alerta. No encontramos semejanzas entre esta vocalización y los sonidos reportados por Cleveland & Snowdon (1982) para titíes cautivos. Por su parte, el pip descrito por Rueda & Zerda (2009) y el pip de Fuentes (2006), presentan características similares a elementos de la vocalización nerviosa.
Identificamos cuatro secuencias diferentes en la vocalización de pelea, dependiendo si hay contacto visual o físico con el grupo contrario. Estas estarían indicando el estado motivacional de cada individuo frente al encuentro intergrupal. Cleveland & Snowdon (1982) presentan silbidos rápidos como una vocalización intragrupo en actividades sociales. Esta es similar a la vocalización de pelea 3 descrita en el presente estudio; sin embargo fue registrada durante un encuentro intergrupal antes del contacto físico entre los machos dominantes de cada uno de ellos. La variación en el contexto en el que es emitido esta vocalización en cautiverio puede deberse a que en los grupos de titíes cautivos nunca hay intromisión a sus territorios.
La vocalización de pelea 4 es similar a la secuencia de sonidos rápidos y agudos (bark bout) descrito por Campbell & Snowdon (2007) para titíes cabeciblancos cautivos; también es similar en forma y características acústicas al trino A reportado por Fuentes (2006) y Fuentes et al. (2013) para el tití gris.
La vocalización de solicitud es la más común en el repertorio de los infantes. Comunica una necesidad por parte de éstos, que debe ser satisfecha por los otros miembros del grupo. No se encontraron sonidos semejantes a éstos en Cleveland & Snowdon (1982) puesto que ellos no reportan vocalizaciones de infantes. Algunos elementos de esta vocalización presentan características similares a chillido B, chillido C y chirr descritos por Rueda & Zerda (2009) para el tití gris. La vocalización de vigilancia, al igual que las vocalizaciones de exploración y nerviosa, es redundante.
La mayor cantidad de sonidos registrados e identificados en este estudio, comparado con los resultados de Cleveland & Snowdon (1982), pueden deberse a que en vida silvestre los titíes tienen la necesidad de coordinar actividades como los desplazamientos, manteniendo la unidad del grupo, así como un espacio mínimo entre los individuos; exploración en busca de alimentos y refugios, defensa del territorio y defensa ante posibles depredadores, necesidades que no están presentes en los grupos mantenidos en cautiverio.
Por otra parte, es posible que existan variaciones en la vocalización de alarma debidas a diferentes tipos de depredadores, como ha sido reportado en otras especies de primates (Seyfarth & Cheney, 1990). También podrían existir variaciones en la vocalización de alimentación relacionadas con la preferencia alimentaria (Elowson et al., 1991). En cautiverio, también se han reportado sonidos relacionados con el comportamiento reproductivo y cuidado parental (Cleveland & Snowdon, 1982), que no fueron registrados en el presente estudio. Por tal razón es necesario continuar con los estudios de la comunicación vocal de este primate en vida silvestre, con el fin de llenar los vacíos de información aún existentes.















