




Recibido: 23 de enero de 2019; Aceptado: 25 de noviembre de 2019
Resumen
Para iniciar un proceso de restauración en áreas disturbadas, es importante seleccionar especies vegetales nativas que aporten diversidad funcional al ecosistema en recuperación y permitan el establecimiento de nuevos individuos hacia etapas sucesionales más avanzadas. Evaluamos 14 rasgos de historia de vida en 20 especies de una zona de transición entre bosque altoandino y páramo en la reserva Aguas Vivas, Soacha, Colombia.
Registramos información de tipos de polinización y dispersión, hábito de crecimiento, altura máxima, tipo y tamaño del fruto, número de semillas por fruto, textura de la hoja, tipo de indumento, área foliar, área foliar específica, nitrógeno foliar, contenido foliar de materia seca y dureza foliar. El análisis de componentes principales mostró que el nitrógeno foliar, la dureza foliar y el área foliar específica fueron los rasgos más informativos para definir asociaciones entre especies. Encontramos tres tipos funcionales que comprenden plantas con diferentes estrategias de adaptación, que se considerarán a la hora de seleccionar grupos de especies vegetales a reintroducir en ambientes modificados. Sin embargo, esta selección también se debe realizar teniendo en cuenta el conocimiento local, la disponibilidad de propágulos, y el estado sucesional de las áreas en recuperación.
Palabras clave
Disturbios, Grupo funcional, Rasgos de historia de vida, Subpáramo, Sucesión vegetal.Abstract
To start a restoration process in disturbed areas, it is important to select plant species that provide functional diversity to the recovering ecosystem and allow the establishment of new individuals towards more advanced stages of succession. We evaluated 14 life history traits in 20 species in a transition zone between high Andean forest and paramo in the Aguas Vivas reserve, Soacha, Colombia. We record information on pollination and dispersal types, growth habit, maximum height, fruit type and size, number of seeds per fruit, leaf texture, leaf area, specific leaf area, leaf nitrogen, leaf dry matter content and leaf hardness. The principal components analysis showed that leaf nitrogen, leaf hardness and specific leaf area were the most informative features to define associations between species. We found three functional types that include plants with different adaptation strategies, which will be considered when selecting plant species to be introduced in modified environments. However, this selection should also be made taking into account local knowledge, availability of propagules, and the successional status of the recovering areas.
Key word
Disturbance, Functional group, Life history traits, Subparamo, Plant succession.Introducción
Los bosques altoandinos se consideran “hotspot” de biodiversidad y encabezan la lista de las áreas más vulnerables a nivel mundial, principalmente por las altas tasas de deforestación y el desarrollo de proyectos que influyen en la transformación acelerada del territorio (Mittermeier & Mittermeier, 1997; Kappelle & Brown, 2001). En Colombia, una buena parte de la diversidad florística se alberga en ecosistemas de alta montaña (Van der Hammen & González, 1960); sin embargo, su pérdida impacta negativamente la riqueza de especies y, por tanto, la diversidad funcional y servicios ecosistémicos que estos brindan (Anderson et al., 2011).
Debido a la oferta hídrica que aportan las áreas rurales del Distrito Capital de Bogotá, es necesario desarrollar estrategias que prioricen la conservación y restauración ecológica de zonas degradadas (Van der Hammen et al., 2008), donde se incluyan especies vegetales con características que reactiven procesos de regeneración natural y posibiliten la restitución de las funciones del sistema y sus características florísticas y estructurales (Naeem et al., 1994; Hooper & Vitousek, 1997). Esta selección se puede simplificar agrupando las especies según sus rasgos funcionales (Castellanos-Castro & Bonilla, 2011; Montenegro & Vargas, 2008; Meli et al., 2014), ya que generalmente las especies adaptadas a ambientes con frecuentes disturbios, ya sean naturales o antrópicos, desarrollan ciertos atributos que contribuyen a la recuperación de áreas disturbadas (Holl, 2002). Además, esta agrupación funcional se considera una herramienta adecuada para el diagnóstico y planteamiento de procesos de restauración ecológica (Kooyman & Rossetto, 2008) y para la conservación en términos de la selección de especies a introducir en ambientes modificados (Castellanos-Castro & Bonilla, 2011).
El objetivo de esta investigación fue la evaluación de rasgos funcionales relacionados a la historia de vida de 20 especies en un sitio de transición entre bosque altoandino y páramo, afectado por la expansión de la frontera agropecuaria, y de esta manera contribuir en la identificación de tipos funcionales importantes en la recuperación de zonas alteradas en ecosistemas de alta montaña.
Materiales y métodos
Área de estudio. La investigación se llevó a cabo en el predio Aguas Vivas, entre los límites de la vereda Hungría en el municipio de Soacha y la vereda Quiba Alta de Bogotá (coordenadas: 4º 28.638’ N, 74º 10.94’ O; 3330 m s. n. m.), durante el periodo comprendido entre los meses de junio de 2013 a junio de 2014. Este predio fue adquirido por la Administración Municipal de Soacha, para preservar la riqueza hídrica de la zona y contribuir con la recuperación de áreas degradadas por uso agropecuario. Cuenta con un área de 97.61 ha de las cuales un 50 % corresponde a coberturas de pastos exóticos donde se evidencia un proceso de sucesión alterada o detenida y el otro porcentaje restante a coberturas naturales de páramo y subpáramo en un mejor estado de conservación.
La reserva es de gran importancia, puesto que aquí se encuentra el nacimiento del río Soacha.
Caracterización vegetal. Se trazaron cinco parcelas de 50 m x 2 m (100 m2), en las que se registró información para todos los individuos de porte arbustivo y arbóreo con alturas (h) mayores a 1 m y diámetro a la altura del pecho (DAP) superior a 1 cm. Se recolectaron datos de h, DAP y cobertura de los individuos. Para la vegetación herbácea se hicieron diez muestreos en cada parcela con ayuda de un cuadrante de 1 m2 que estaba subdividido en 100 cuadrados de 10 cm2 (para facilitar la estimación de la cobertura), y en cada muestreo se midió la altura promedio con ayuda de un flexómetro y el porcentaje de cobertura de todas las especies con altura menor a 1 m. Se recolectó el material vegetal para su posterior determinación y se depositó en el herbario del Jardín Botánico José Celestino Mutis (JBB; http://colecciones.jbb.gov.co/herbario).
Selección de las especies. Se escogieron las especies dominantes según el Índice de Valor de Importancia (IVI), el cual se calcula para cada especie a partir de la suma de la abundancia relativa, la frecuencia relativa y la dominancia relativa (Curtis & McIntosh, 1951).
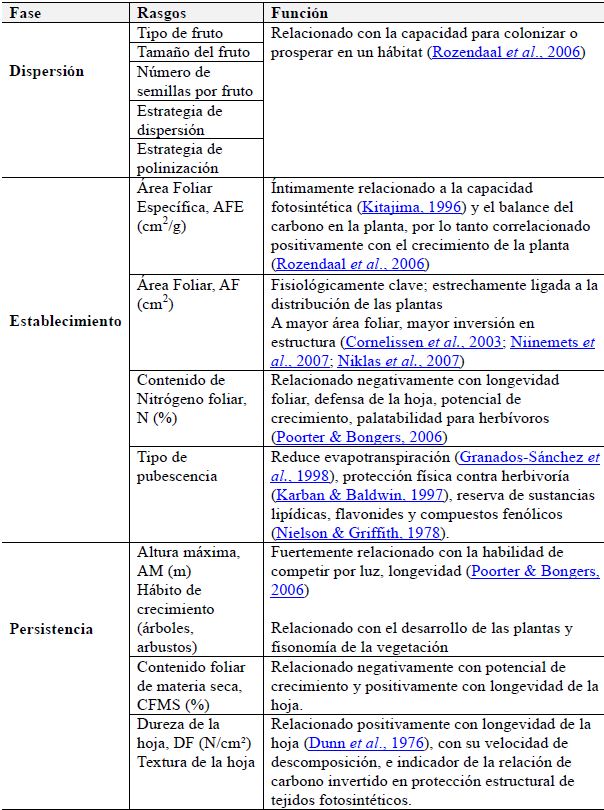
Selección de rasgos funcionales. Los mejores rasgos son aquellos que desde el punto de vista ecológico ayudan a superar las limitaciones bióticas y abióticas a la regeneración natural (disponibilidad de propágulos, condiciones climáticas, oferta hídrica y de nutrientes, entre otros) y pueden ser medidos en un menor tiempo, a un menor costo y más fácilmente, para un número representativo de individuos dentro de una población (Cornelissen et al., 2003). Con base en esto, se seleccionaron algunos rasgos asociados a la fase de dispersión, establecimiento y persistencia en especies vegetales, según lo propuesto por Weiher et al. (1999), Lavorel et al. (1998), Díaz & Cabido (1997) (Tabla 1).
Tabla 1: Rasgos funcionales de plantas asociados a la fase de dispersión, establecimiento y persistencia
FASE
RASGOS
FUNCIÓN
Tipo de fruto
Relacionado con la capacidad para colonizar o prosperar en un hábitat (Rozendaal et al., 2006)
Tamaño del fruto
Dispersión
Número de semillas por fruto
Estrategia de dispersión
Estrategia de polinización
Área Foliar Específica, AFE (cm2.g-1)
Íntimamente relacionado a la capacidad fotosintética (Kitajima, 1996) y el balance del carbono en la planta, por lo tanto correlacionado positivamente con el crecimiento de la planta (Rozendaal et al., 2006)
Establecimiento
Área Foliar, AF (cm2)
Fisiológicamente clave; estrechamente ligada a la distribución de las plantas A mayor área foliar, mayor inversión en estructura (Cornelissen et al., 2003; Niinemets et al., 2007; Niklas et al., 2007)
Contenido de Nitrógeno foliar, N (%)
Relacionado negativamente con longevidad foliar, defensa de la hoja, potencial de crecimiento, palatabilidad para herbívoros (Poorter & Bongers, 2006)
Tipo de pubescencia
Reduce evapotranspiración (Granados-Sánchez et al., 1998), protección física contra herbivoría (Karban & Baldwin, 1997), reserva de sustancias lipídicas, flavonides y compuestos fenólicos (Nielson & Griffith, 1978).
Persistencia
Altura máxima, AM (m) Hábito de crecimiento (árboles, arbustos)
Fuertemente relacionado con la habilidad de competir por luz, longevidad (Poorter & Bongers, 2006) Relacionado con el desarrollo de las plantas y fisonomía de la vegetación
Contenido foliar de materia seca, CFMS (%)
Relacionado negativamente con potencial de crecimiento y positivamente con longevidad de la hoja.
Dureza de la hoja, DF (N.cm-²) Textura de la hoja
Relacionado positivamente con longevidad de la hoja (Dunn et al., 1976), con su velocidad de descomposición, e indicador de la relación de carbono invertido en protección estructural de tejidos fotosintéticos.
Obtención de los datos. Para la evaluación de los rasgos foliares se seleccionaron cinco individuos por especie. Por cada individuo adulto se recolectó una muestra aleatoria en promedio de cinco hojas maduras completamente desarrolladas, sin evidencia de daños por patógenos ni herbivoría, siguiendo el protocolo para la toma de muestras foliares de Garnier et al. (2001).
El área foliar (AF) se determinó escaneando las hojas frescas a través de un escáner marca HP psc 1310. Después de esto, las hojas se secaron durante 48 horas en un horno a 70 °C y se pesaron, para luego determinar el área foliar específica (AFE) (promedio del área foliar/ promedio del peso seco) por medio del software libre ImageJ (Rasband, 2007).
El contenido foliar de materia seca (CFMS) se realizó a través de la eliminación del agua libre por medio del calor, seguida por la determinación del peso del residuo (Batteman, 1970); la cuantificación del nitrógeno foliar (N) se hizo siguiendo la metodología de Kjeldahl (AOAC, 2000). Este procedimiento se llevó a cabo en el laboratorio de Bromatología del Jardín Botánico José Celestino Mutis.
Para la caracterización de los tipos de indumento, se recolectaron hojas maduras, que tuvieran un área foliar similar. Se observó el limbo de cada hoja por las caras adaxial y abaxial, y se identificó el tipo de indumento de cada una de las especies analizadas con la ayuda de un estereoscopio marca Nikon SMZ-1000. Para la identificación de los indumentos se utilizó el glosario Ilustrado de términos botánicos de Harris & Harris (1994). Para medir la dureza foliar (DF), cada hoja fue sostenida con dos pinzas botánicas por el ápice y la base del limbo, y con la ayuda de un penetrómetro marca EXPOTECH referencia SHC 280, se calculó la fuerza necesaria para atravesar la zona central al lado derecho de la vena media de cada una de las hojas, calculada en newtons por centímetro cuadrado (N.cm-²), método descrito por Hendry & Grime (1993). Con los datos obtenidos se calculó el promedio y la desviación estándar para obtener los valores de DF de cada especie. La textura de la hoja se determinó con ayuda del Glosario Botánico Ilustrado (Moreno, 1984).
Para evaluar los rasgos reproductivos, se recolectaron en promedio quince frutos a partir de cinco individuos de la misma especie; estos debían estar en lo posible maduros en el caso de bayas, drupas o legumbres y secos en el caso de los aquenios. En fresco, se tomaron medidas de longitud (mm) y diámetro (mm) para cada uno de los frutos con un calibrador con una precisión de 0.001. Los valores obtenidos de cada fruto se promediaron para cada una de las especies. También se registró el número de semillas/fruto, las cuales se contaban en su totalidad, exceptuando los frutos de las especies que presentaran un número mayor a 50 semillas, utilizando un estereoscopio marca Nikon SMZ-1000. El tipo de fruto se determinó con base en su aspecto morfológico y la forma de crecimiento con base en la clasificación de Moreno (1984).
La información sobre estrategia de dispersión y polinización se tomó de la literatura y de datos en el campo, teniendo en cuenta la forma y estructura de la semilla y de la flor. La altura máxima se obtuvo de los levantamientos de información vegetal que se tomaron en el campo.
Identificación de tipos funcionales. Se utilizó el enfoque de tipos funcionales de plantas (TFP), el cual agrupa a las especies con funcionamiento similar, independientemente de su filogenia (Gitay et al., 1999; Kooyman & Rosetto, 2008; Delgado, 2012). Las plantas que conforman los TFP exhiben respuestas similares frente a las condiciones ambientales y también producen efectos similares en los procesos ecosistémicos (Díaz & Cabido, 1997).
Análisis de datos. A partir de los valores obtenidos para cada una de las variables cuantitativas se realizó la prueba de Shapiro Wilks para establecer si los datos siguen una distribución Gaussiana. Con ayuda del software PAST 7.0 (Hammer & Harper, 2005) se realizó un análisis de los componentes principales (ACP), para la selección de los parámetros que más refuerzan el agrupamiento. El análisis busca agrupar las especies en función de las variables medidas y encontrar de manera intuitiva las causas del agrupamiento. Para el ACP solo se utilizaron variables cuantitativas continuas de rasgos foliares (AF, AFE, N, CFMS y DF), asumiendo que los datos deben cumplir con el supuesto de normalidad.
Se realizó un Análisis de Conglomerados (clúster) con el programa Statsoft, para la determinación de los tipos funcionales de plantas, TFP. Una vez realizado dicho análisis y agrupadas las especies a partir de la totalidad de rasgos funcionales evaluados, se conformaron y caracterizaron los tipos funcionales resultantes. Adicionalmente, para confirmar o rechazar la hipótesis de que los grupos de plantas resultantes del análisis clúster son distintos, se realizó un análisis discriminante (AD) a partir de la prueba de T-cuadrado de Hotelling, que permite la clasificación de dichos grupos con relación a los valores encontrados en los rasgos foliares en conjunto y las posibles correlaciones presentes en las mismas. Dados dos grupos de datos multivariados, se comparan las medias de las variables clasificadoras a través de los grupos obtenidos previamente; a partir de los valores de la función discriminante se establecen cuáles son los rasgos que más discriminan (separan) las especies en cada grupo (Hammer & Harper, 2005). Finalmente, se realizó un análisis de correlación de Pearson entre los rasgos foliares cuantitativos (AFE, AF, CFMS, N, DF) para establecer el grado de correlación entre las cinco variables.
Resultados
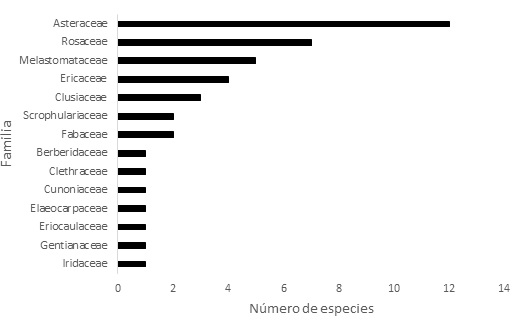
En el inventario de vegetación se encontraron 50 especies distribuidas en 22 familias. Las familias más representativas fueron Asteraceae (12 especies), Rosaceae (7 especies) y Melastomataceae (5 especies; Figura 1).
Figura 1: Número de especies de plantas por familia en el predio Aguas Vivas, Soacha-Bogotá, Colombia.
A partir del IVI, las especies con mayor dominancia fueron Bucquetia glutinosa, Pentacalia pulchella, Miconia ligustrina, Ageratina tinifolia, Miconia elaeoides y Macleania rupestris.
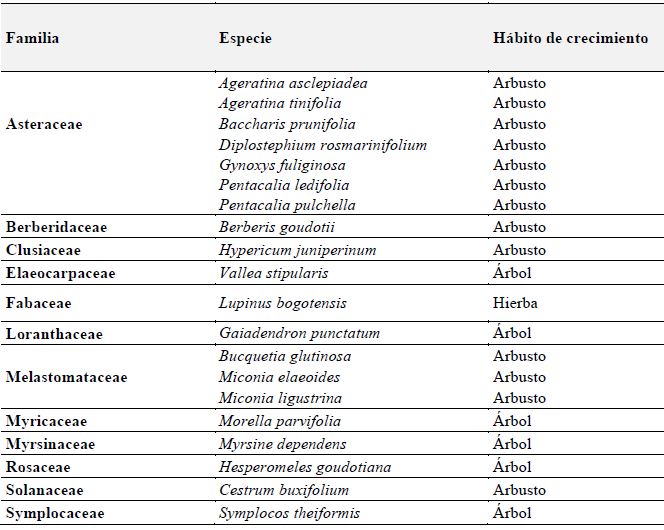
Para la evaluación de los rasgos de historia de vida, se seleccionaron en su mayoría especies del estrato arbóreo-arbustivo (Tabla 2), para las cuales se tuvieron en cuenta la dominancia, observaciones en campo y su utilización en procesos de restauración ecológica adelantados en el Jardín Botánico de Bogotá.
Tabla 2: Especies de plantas seleccionadas para la evaluación de los rasgos funcionales en el predio Aguas Vivas, Soacha-Bogotá, Colombia
Familia
Especie
Hábito de crecimiento
Ageratina asclepiadea
Arbusto
Ageratina tinifolia
Arbusto
Asteraceae
Baccharis prunifolia
Arbusto
Diplostephium rosmarinifolium
Arbusto
Gynoxys fuliginosa
Arbusto
Pentacalia ledifolia
Arbusto
Pentacalia pulchella
Arbusto
Berberidaceae
Berberis goudotii
Arbusto
Clusiaceae
Hypericum juniperinum
Arbusto
Elaeocarpaceae
Vallea stipularis
Árbol
Fabaceae
Lupinus bogotensis
Hierba
Loranthaceae
Gaiadendron punctatum
Árbol
Bucquetia glutinosa
Arbusto
Melastomataceae
Miconia elaeoides
Arbusto
Miconia ligustrina
Arbusto
Myricaceae
Morella parvifolia
Árbol
Myrsinaceae
Myrsine dependens
Árbol
Rosaceae
Hesperomeles goudotiana
Árbol
Solanaceae
Cestrum buxifolium
Arbusto
Symplocaceae
Symplocos theiformis
Árbol
Rasgos funcionales.
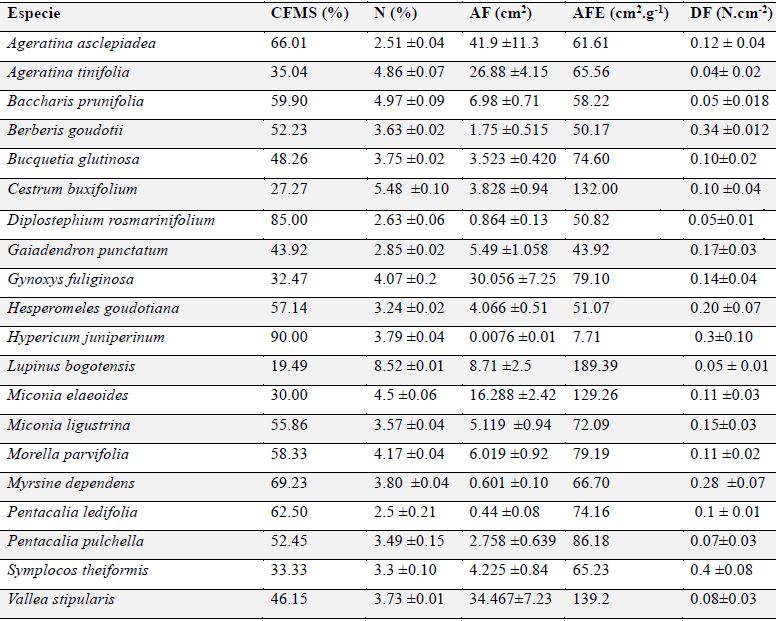
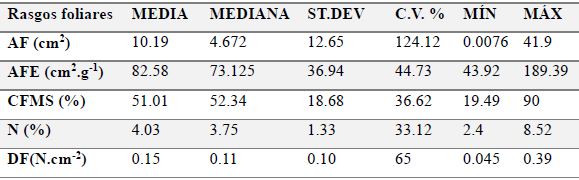
Para los rasgos foliares se determinó el peso seco, peso fresco, CFMS, N, AF, AFE y DF para las 20 especies (Tabla 3). La prueba de normalidad multivariante estableció una distribución normal de los valores promedio provenientes de las variables AF, AFE, N, CFMS y DF (α> 0.05 g.l.= 35 n= 100).
Tabla 3: Rasgos foliares evaluados en las 20 especies de plantas seleccionadas en el predio Aguas Vivas, Soacha-Bogotá, Colombia.
Especie
CFMS (%)
N (%)
AF (cm2)
AFE (cm2.g-1)
DF (N.cm-2)
Ageratina asclepiadea
66.01
2.51 ±0.04
41.9 ±11.3
61.61
0.12 ± 0.04
Ageratina tinifolia
35.04
4.86 ±0.07
26.88 ±4.15
65.56
0.04± 0.02
Baccharis prunifolia
59.90
4.97 ±0.09
6.98 ±0.71
58.22
0.05 ±0.018
Berberis goudotii
52.23
3.63 ±0.02
1.75 ±0.515
50.17
0.34 ±0.012
Bucquetia glutinosa
48.26
3.75 ±0.02
3.523 ±0.420
74.60
0.10±0.02
Cestrum buxifolium
27.27
5.48 ±0.10
3.828 ±0.94
132.00
0.10 ±0.04
Diplostephium rosmarinifolium
85.00
2.63 ±0.06
0.864 ±0.13
50.82
0.05±0.01
Gaiadendron punctatum
43.92
2.85 ±0.02
5.49 ±1.058
43.92
0.17±0.03
Gynoxys fuliginosa
32.47
4.07 ±0.2
30.056 ±7.25
79.10
0.14±0.04
Hesperomeles goudotiana
57.14
3.24 ±0.02
4.066 ±0.51
51.07
0.20 ±0.07
Hypericum juniperinum
90.00
3.79 ±0.04
0.0076 ±0.01
7.71
0.3±0.10
Lupinus bogotensis
19.49
8.52 ±0.01
8.71 ±2.5
189.39
0.05 ± 0.01
Miconia elaeoides
30.00
4.5 ±0.06
16.288 ±2.42
129.26
0.11 ±0.03
Miconia ligustrina
55.86
3.57 ±0.04
5.119 ±0.94
72.09
0.15±0.03
Morella parvifolia
58.33
4.17 ±0.04
6.019 ±0.92
79.19
0.11 ±0.02
Myrsine dependens
69.23
3.80 ±0.04
0.601 ±0.10
66.70
0.28 ±0.07
Pentacalia ledifolia
62.50
2.5 ±0.21
0.44 ±0.08
74.16
0.1 ± 0.01
Pentacalia pulchella
52.45
3.49 ±0.15
2.758 ±0.639
86.18
0.07±0.03
Symplocos theiformis
33.33
3.3 ±0.10
4.225 ±0.84
65.23
0.4 ±0.08
Vallea stipularis
46.15
3.73 ±0.01
34.467±7.23
139.2
0.08±0.03
El rasgo foliar más heterogéneo fue el AF (124.12%) que se debe principalmente a la variabilidad aportada por las especies Ageratina asclepiadea y Vallea stipularis. Los demás rasgos tuvieron una mayor homogeneidad, presentando coeficientes de variación menores al 45%, exceptuando la dureza foliar (Tabla 4).
Tabla 4: Estadística descriptiva para los rasgos foliares de 20 especies de plantas seleccionadas en el predio Aguas Vivas, Soacha-Bogotá, Colombia. ST.DEV, desviación estándar; C.V., coeficiente de variación; MIN, valor mínimo; MÁX, valor máximo.
Rasgos foliares
MEDIA
MEDIANA
ST.DEV
C.V. %
MÍN
MÁX
AF (cm2)
10.19
4.672
12.65
124.12
0.0076
41.9
AFE (cm2.g-1)
82.58
73.125
36.94
44.73
43.92
189.39
CFMS (%)
51.01
52.34
18.68
36.62
19.49
90
N (%)
4.03
3.75
1.33
33.12
2.4
8.52
DF(N.cm-2)
0.15
0.11
0.10
65
0.045
0.39
Los tipos de indumento en las hojas fueron glabro (Bucquetia glutinosa, Miconia ligustrina, Pentacalia pulchella, Gaiadendron punctatum, Cestrum buxifolium, Vallea stipularis, Hypericum juniperinum, Symplocos theiformis, Ageratina tinifolia, Miconia elaeoides, Berberis goudotii), manicado (Diplostephium rosmarinifolium, Morella parvifolia, Hesperomeles goudotiana), piloso (Baccharis prunifolia, Lupinus bogotensis), panoso (Myrsine dependens), lanado ( Gynoxys fuliginosa) y seríceo (Pentacalia ledifolia). Las especies con mayor producción de semillas fueron las pertenecientes a las familias Melastomataceae y Asteraceae, y las de menor producción: Gaiadendron punctatum, Morella parvifolia, Myrsine dependens y Symplocos theiformis. El 35 % de las especies presentaron frutos de tipo aquenio (Pentacalia pulchella, Ageratina asclepiadea, Diplostephium rosmarinifolium, Gynoxys fuliginosa, Baccharis prunifolia, Ageratina tinifolia, Pentacalia ledifolia), seguidas por bayas (Miconia ligustrina, Cestrum buxifolium, Miconia elaeoides, Berberis goudotii) y drupas (Gaiadendron punctatum, Morella parvifolia, Myrsine dependens, Symplocos theiformis) con 20 % cada una; las demás especies presentan cápsulas (Bucquetia glutinosa, Vallea stipularis, Hypericum juniperinum), legumbres (Lupinus bogotensis) y pomos (Hesperomeles goudotiana). Las estrategias de dispersión que se encontraron fueron balocoria, barocoria, zoocoria y anemocoria.
La forma de crecimiento que predominó fue la de tipo arbustivo (70 % de las especies) y las especies que registraron mayor altura fueron Gaiadendrom punctatum, Vallea stipularis y Miconia ligustrina con valores de 8, 7 y 6 metros, respectivamente.
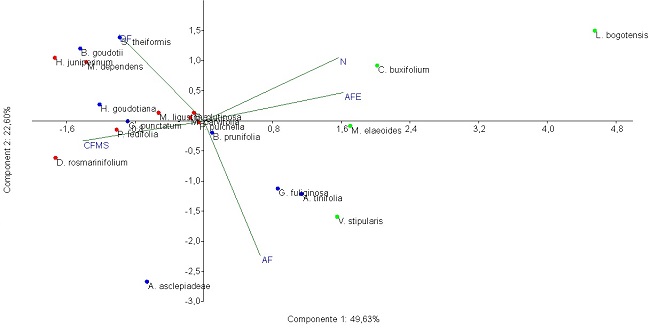
Análisis multivariados. En el análisis de componentes principales, los dos primeros componentes explicaron el 72.23 % de la variabilidad total (49.63 % y 22.6 %, respectivamente). En ésta, los valores de AFE y N se asociaron negativamente con el CFMS, de igual manera el AF tuvo una relación negativa con la DF (Figura 2).
Figura 2: Diagrama de ordenación de las especies de plantas seleccionadas en el predio Aguas Vivas, Soacha-Bogotá, Colombia, en los dos primeros ejes establecidos en el ACP. AF, área foliar; AFE, área foliar específica; CFMS, contenido foliar de materia seca; DF, dureza foliar; N, contenido de nitrógeno. Puntos rojos, TFP1; puntos azules, TFP2; puntos verdes, TFP3. Valor propio eje 1= 2.48; valor propio eje 2= 1.12.
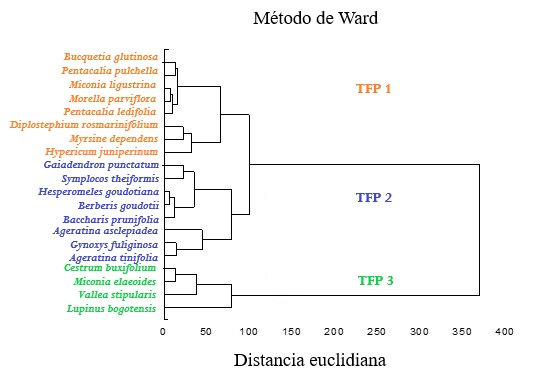
A través del análisis clúster, se diferenciaron tres tipos funcionales de acuerdo a sus similitudes descritas en los rasgos de historia de vida (Figura 3).
Figura 3: Análisis de conglomerados de plantas seleccionadas en el predio Aguas Vivas, Soacha-Bogotá, Colombia, basado en rasgos categóricos (polinización, dispersión, tipo de fruto, forma de crecimiento, textura de la hoja, tipo de indumento), categóricos cuantitativos (tamaño del fruto, número de semillas por fruto) y cuantitativos (AF, AFE, N, CFMS, DF, AM).
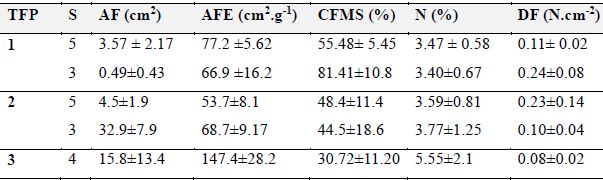
El TFP1 lo conformaron ocho especies que presentaron en general los valores más bajos de nitrógeno foliar. Sin embargo, este grupo se dividió en dos subclados: el primero con cinco especies (Bucquetia glutinosa, Pentacalia pulchella, Miconia ligustrina, Morella parvifolia y Pentacalia ledifolia) que registraron en promedio valores de AF de 3.57 ± 2.17 cm2 y CFMS de 55.48 % ± 5.45 %. El segundo subclado con tres especies (Diplostephium rosmarinifolium, Myrsine dependens e Hypericum juniperinum) que presentaron las hojas más pequeñas con valores promedio de AF de 0.49 ± 0.43 cm2 y el más alto contenido de materia seca con 81.41 ± 10.8 %.
El TFP 2 está conformado por ocho especies, todas con estrategia de dispersión por ornitocoria excepto Baccharis prunifolia y con valores intermedios de CFMS y de N. En este tipo funcional se presentan dos subclados, el primero lo conforman las especies Gaiadendrom punctatum, Symplocos theiformis, Hesperomeles goudotiana, Berberis goudotii y Baccharis prunifolia, que registraron el menor valor de AFE con 53.7 ± 8.1 cm2.g-1 y otro subclado conformado por Ageratina asclepiadea, Gynoxys fuliginosa y Ageratina tinifolia, especies cuyas hojas fueron las de mayor AF, con 32.9 ± 7.9 cm2.
En el TFP 3 están las especies Cestrum buxifolium, Miconia elaeoides, Vallea stipularis y Lupinus bogotensis. Todas con hojas de textura membranosa. En este grupo están las especies con mayores valores de contenido de N y AFE y los valores más bajos de CFMS y DF (Tabla 5).
Tabla 5: Promedio y desviación estándar para los rasgos foliares de cada tipo funcional de plantas (TFP) en el predio Aguas Vivas, Soacha-Bogotá, Colombia. AF, área foliar; AFE, área foliar específica; CFMS, contenido foliar de materia seca; DF, dureza foliar; N, contenido de nitrógeno; S, número de especies.
TFP
S
AF (cm2)
AFE (cm2.g-1)
CFMS (%)
N (%)
DF (N.cm-2)
1
5
3.57 ± 2.17
77.2 ±5.62
55.48± 5.45
3.47 ± 0.58
0.11± 0.02
3
0.49±0.43
66.9 ±16.2
81.41±10.8
3.40±0.67
0.24±0.08
2
5
4.5±1.9
53.7±8.1
48.4±11.4
3.59±0.81
0.23±0.14
3
32.9±7.9
68.7±9.17
44.5±18.6
3.77±1.25
0.10±0.04
3
4
15.8±13.4
147.4±28.2
30.72±11.20
5.55±2.1
0.08±0.02
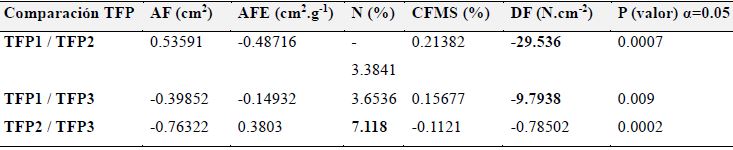
La prueba T cuadrado de Hotteling mostró diferencias significativas entre los 3 TFP (α<0.05), lo cual indica que existe un agrupamiento de las especies a partir de los valores obtenidos para los diferentes rasgos foliares. En cuanto al análisis discriminante, se encontró que la DF es el rasgo de mayor discriminación entre el TFP1 y TFP2 y entre el TFP1 y TFP3, mientras que para el TFP2 y TFP3 es el N quien diferencia estos grupos (Tabla 6).
Tabla 6: Valores comparativos de la función discriminante para los tres tipos funcionales de plantas (TFP) en el predio Aguas Vivas, Soacha-Bogotá, Colombia, con base en 5 rasgos foliares. AF, área foliar; AFE, área foliar específica; CFMS, contenido foliar de materia seca; DF, dureza foliar; N, contenido de nitrógeno; S, número de especies. Nivel de confianza, 95 %.
Comparación TFP
AF (cm2)
AFE (cm2.g-1)
N (%)
CFMS (%)
DF (N.cm-2)
P (valor) α=0.05
TFP1 / TFP2
0.53591
-0.48716
-3.3841
0.21382
-29.536
0.0007
TFP1 / TFP3
-0.39852
-0.14932
3.6536
0.15677
-9.7938
0.009
TFP2 / TFP3
-0.76322
0.3803
7.118
-0.1121
-0.78502
0.0002
Hubo una correlación directa (0.76) entre el AFE y N, que son dos rasgos relacionados con la capacidad de fotosíntesis (Kitajima, 1996). Por otro lado, el N y CFMS se relacionaron negativamente (-0.56). Esto se evidenció en Lupinus bogotensis y Cestrum buxifolium, que fueron las plantas que presentaron mayor contenido de N y el menor contenido de materia seca en la hoja. El CFMS se relacionó negativamente con el AFE (-0.53), ya que un mayor peso seco da menores valores de AFE (Bermeo, 2010). Esto se representa muy bien en Hypericum juniperinum, que fue la especie con mayor CFMS (90 %) y menor AFE (0.076 cm2.g-1). La DF se relacionó negativamente con el AF (-0.36), siendo Myrsine dependens, Berberis goudotii y Symplocos theiformis las especies de mayor dureza foliar, pero que presentaban hojas pequeñas.
El AFE y el AF se correlacionan positivamente; esto se puede deber a que estas dos variables según Kitajima (1996) están correlacionadas con la tasa fotosintética y con el contenido de nitrógeno foliar por unidad de peso foliar. Otra correlación positiva se da entre AF-N y una correlación negativa entre CFMS-AF.
Discusión
En el TFP1 se encuentran las especies con mayor DF y CFMS y menor N, como Diplostephium rosmarinifolium, Myrsine dependens e Hypericum juniperinum. La DF, puede estar correlacionada con una defensa física más eficaz, puesto que las hojas duras son más difíciles de cortar y digerir por los herbívoros (Barone & Coley, 2002). Por otro lado, el N puede incidir sobre la intensidad de la herbivoría, sin embargo, esta última también dependerá de la disponibilidad de nutrientes y la edad de las hojas, es decir, cuanto más joven la hoja, mayor será el daño por herbivoría (Coley, 1990)
El TFP 2 presenta especies con valores bajos de AFE e intermedios de N. Estas especies podrían encajar dentro de la estrategia conservativa, que se caracteriza por ser de sucesión tardía y crecimiento lento (Hobbie, 1992), ya que son especies con AFE baja, la cual se relaciona con la tasa de crecimiento de las plantas (Cornelissen et al, 2003), hojas duras y bajo contenido de nitrógeno (Kühner & Kleyer, 2008). En un proceso de restauración ecológica, estas especies del TPF 2 podrían plantarse en una etapa posterior al establecimiento de las especies tolerantes a la exposición solar, luego de generarse una cobertura que permita hacer enriquecimientos (Knowles & Parrotta, 1995).
En el TFP 2, las especies con menor AFE fueron Gaiadendron punctatum (43.92 cm2.g-1), Berberis goudotii (50.17 cm2.g-1) y Hesperomeles goudotiana (51.07 cm2.g-1), lo cual se asocia con hojas más longevas y con más ventajas en hábitats con escasez de nutrientes y agua (Cornelissen et al., 1996).
Tanto en el TFP1 como en el TFP2 se registraron altos valores de DF y CFMS, rasgos presentes en especies que dominan altitudes intermedias (Garnica & Saldarriaga, 2015), como se reportó en el complejo de Páramo Sumapaz - Cruz Verde y se establecen en áreas de conectividad entre el bosque altoandino y el páramo. Especies con estos rasgos podrían considerarse de crecimiento lento, y de acuerdo a la hipótesis de disponibilidad de recursos de Coley et al. (1985), estas especies invierten más en defensa contra los herbívoros, puesto que están adaptadas a vivir en áreas con pobres condiciones edáficas.
En el TFP 3 están las especies con mayor AFE, como Lupinus bogotensis (189.39 cm2.g-1), Vallea stipularis (139.2 cm2.g-1), Cestrum buxifolium (132 cm2.g-1) y Miconia elaeoides (129.26 cm2.g-1). El AFE se relaciona directamente con la capacidad para adquirir energía solar y dióxido de carbono, y por tanto, especies con estas características obtienen una mayor ganancia de carbono, reflejada en altas tasas de crecimiento y mayor capacidad competitiva (Wright et al., 2004). También el AFE podría relacionarse con la longevidad foliar, siendo las las hojas de especies con mayor AFE menos longevas, pues especies con esta característica tienden a invertir una menor cantidad de energía en la defensa de las hojas, pero a cambio de tasas metabólicas más altas (Cornelissen et al., 2003). En este grupo las especies tienen hojas de textura membranosa, son las que presentan mayores valores de contenido N y los valores más bajos de CFMS, que según Grime et al. (1997) podrían corresponder a especies de estrategia adquisitiva, que se caracterizan por ser colonizadoras, con tasas de crecimiento rápido, de hojas delgadas, suaves, con alta AFE y bajo CFMS. Según Weiher et al. (1999), una tasa de crecimiento rápido se ve representada en áreas foliares grandes, debido a que está relacionada con la estrategia de aprovechamiento de luz (Garnier et al., 2001), y por lo tanto, con una mayor fotosíntesis. Este atributo es importante para la restauración ecológica, ya que las especies con altas tasas de crecimiento tienen la capacidad de acumular biomasa de manera más rápida, y pueden actuar como facilitadoras para el establecimiento de otras especies que requieren determinados microclimas para adaptarse adecuadamente (Gómez & Vargas, 2011) o como competidoras frente al avance de especies exóticas invasoras en ecosistemas altoandinos, como sucede con Lupinus bogotensis, usada para el control de Ulex europaeus (Vargas et al, 2009). Por otro lado, el alto contenido de nitrógeno en las hojas es una característica que puede emplearse para iniciar procesos de restauración en suelos degradados y susceptibles a la erosión, dado que el N está relacionado con la tasa de descomposición de la hojarasca la cual aporta rápidamente materia orgánica al suelo (León et al., 2008).
Se ha sugerido que las especies de hojas coriáceas, de alta dureza foliar y bajo contenido de nitrógeno, se desarrollan favorablemente en hábitats donde los suelos son pobres (Loveless, 1962). Tal puede ser el caso de Diplostephium rosmarinifolium, Myrsine dependens, Hypericum juniperinum, Gaiadendrom punctatum, Symplocos theiformis, Hesperomeles goudotiana, Berberis goudotii y Baccharis prunifolia¸ las cuales presentaron hojas coriáceas, de mayor dureza foliar que las demás especies evaluadas. Este tipo de especies tienden a presentar bajas tasas de crecimiento y por lo tanto a requerir menos nutrientes, lo que se ve reflejado en hojarasca con deficiencia de nutrientes minerales, que se liberan lentamente al bosque (Valladares, 2008).
Las especies de hojas coriáceas presentan las paredes celulares engrosadas y espesas cutículas protectoras, que puede ser una ventaja cuando la planta se encuentra sometida a estrés hídrico, ya que se evita la pérdida de agua (Larcher, 2003); por tal razón, este atributo se podría considerar para la selección de especies que se adapten a condiciones de sequía.
En cuanto a la producción de semillas, algunos autores (Janzen, 1970; Hubbell, 1980; McCanny, 1985) consideran que existe una relación inversa entre semillas producidas y supervivencia de las plántulas. Es decir, plantas que producen semillas pequeñas tienen como estrategia de propagación la producción de abundantes semillas para aumentar la probabilidad de supervivencia de algunas plántulas, como puede ocurrir en nuestra área de estudio con las especies de Melastomatacea y Asteraceae. Este carácter es útil para la restauración ecológica, ya que la producción de semillas también puede estar relacionada con la cantidad de dispersores que arriban al lugar, que podría incrementar el grado de conectividad en las zonas a restaurar (Albuquerque et al., 2013).
Por el contrario, en especies como Gaiadendron punctatum, Morella parvifolia, Myrsine dependens y Symplocos theiformis se pueden producir pocas semillas, pero dotadas individualmente de mayor cantidad de reservas (semillas de 5-10 mm) que darán lugar a plántulas de mayor tamaño y con mayor probabilidad de sobrevivir (Coomes & Grubb, 2003). Sin embargo, un tamaño pequeño de semilla también puede permitir que las especies se reproduzcan más rápidamente, inicien procesos sucesionales y colonicen sitios antes que otras especies, mientras que semillas pesadas de tamaño grande tienden a tener un mejor establecimiento cuando compiten con sus vecinos (Weiher et al., 1999).
Conclusión
Los TFP estuvieron asociados principalmente a variables foliares como el contenido de N, DF y el AFE, más que a la altura de la planta, tipos de fruto, tipo de indumento o hábito de crecimiento. Los rasgos foliares resultaron ser muy informativos a la hora de definir diferentes asociaciones, pues las especies que se identificaron como de rápido crecimiento registraron altos valores de AFE, N, textura de hoja membranosa y bajos contenidos de CFMS tal como se registró en el TFP3 compuesto por Cestrum buxifolium Miconia elaeoides, Vallea stipularis y Lupinus bogotensis. El TFP1 lo conformaron especies con menor AF y mayor CFMS y DF, correlacionándose positivamente entre sí estas dos últimas variables, mientras que el TFP2 se diferenció de los demás tipos por presentar las especies con mayor AF, destacándose el aporte de la familia Asteraceae, pues representaban el 50% de las especies del grupo. Son necesarios más estudios para evaluar diferentes rasgos funcionales que contribuyan a la recuperación de los ecosistemas en respuesta a las perturbaciones naturales o antrópicas. Características como producción de hojarasca, rasgos radiculares y de la madera, palatabilidad de las hojas, entre otros, se deben considerar a la hora de escoger asociaciones de plantas a incorporar en los diseños; sin embargo, esta selección también se hace teniendo en cuenta las condiciones del sitio a restaurar, el estado de conservación de las especies, las características fisionómico-estructurales de la vegetación presente y los objetivos y metas del proyecto a alcanzar.















